![]()
![]()
![]()
J. Klein,1 Zh. Xinquan2 and J. Harte1
1. University of California, Department of Environmental Science, Policy & Management, Division of Ecosystem Science, 151 Hilgard Hall #3110, Berkeley, CA
94720–3110 USA
2. Northwest Plateau Institute of Biology, Chinese Academy of Sciences, Xining 810001, Qinghai, P.R. China
This study investigates how two cited causes of rangeland degradation—grazing and climate change—affect key ecological characteristics of the rangelands on the north-eastern Tibetan Plateau. In August 1997, we established experimental plots in two habitats: 1) a summer-grazed shrubland; and 2) a winter-grazed meadow. Within each habitat, we identified sites with low and high grazing intensity histories, within which we established a complete factorial experimental design. The treatments are: 1) simulated warming using open top chambers (OTC); 2) simulated grazing through clipping (G); and 3) combined warming and defoliation (OTC × G). In this paper, we present 1999 growing-season data on habitat, grazing history and treatment effects on vegetative aboveground biomass. Combining all sites, the growing-season-averaged air temperatures in the OTC and OTC × G plots were 1.28°C and 1.33°C warmer than the controls, respectively. Moreover, the chambered plots (OTC and OTC × G treatments combined) resulted in a 12% decline (P = 0.07) in total aboveground biomass. However, different trends emerge when the sites are grouped by grazing history. In the low grazing history sites, the defoliated plots (G and OTC × G treatments combined) had approximately 17% more total biomass compared to the non-defoliated plots. Conversely, in the high grazing history sites, the chambered plots had approximately 12% less biomass as compared to the non-chambered plots. Furthermore, the shrub sites were more responsive to the treatments as compared to the meadow sites. To relate these biomass changes to rangeland quality, we evaluate the compositional changes of different vegetative functional groups in response to our experimental manipulations.
Keywords: Climate warming, grazing, rangelands, Tibetan Plateau, vegetative biomass
The Tibetan Plateau is widely regarded as a region where the traditional pastoral system, which has developed over millennia, still flourishes. Over 70% of the vegetation that covers 2.5 million km2 of the Plateau is classified as 'rangeland' vegetation. In the relatively mesic and productive north-eastern portion of the Tibetan Plateau, the primary habitat types are alpine meadow and shrub vegetation; these habitats comprise approximately 35% of the total rangeland area occupied by the Tibetan Plateau (Zhao and Zhou 1999). The vegetation in this region provides relatively favourable forage for yak and sheep, the pastoralists' primary livestock. However, as in most parts of the world, this region is facing a suite of perturbations—changes that may profoundly affect the natural vegetation that constitutes the foundation of this pastoral production system. The driving forces behind these modifications to the rangelands are varied—some may be attributed to natural phenomena and cycles; some result from human activities far beyond the margin of the Tibetan Plateau; and some of these changes may result from dynamics within the region itself.
One phenomenon that may be affecting the Plateau grasslands is a warming and drying trend. Direct field measurements (French and Wang 1994), ice core data (Thompson et al. 1989; Yao et al. 1997), and interviews with pastoralists over 25 years old living in the research area (unpublished data), all provide evidence of a warming trend on the north-eastern Tibetan Plateau. Concurrent with these climatic changes, over the past 10–20 years there have been changes to the pastoral land use dynamic due to agricultural development, land tenure and sedentarisation policies (Cincotta et al. 1992; Miller 1999b).
Researchers (Yang 1992; Miller 1999b; Limbach et al. 2000; Ma et al. 2000) and pastoralists have observed rangeland degradation in recent years on the Tibetan Plateau. Some researchers attribute this to overgrazing (Ma et al. 2000); others suggest that climate change may be responsible for these rangeland dynamics (Miller 1999b). The purpose of this paper is to present data from a study that is explicitly investigating how two cited causes of rangeland degradation—grazing and climate change—affect key ecological characteristics of the rangelands on the north-eastern Tibetan Plateau. Through controlled experiments, we seek to obtain a detailed understanding of climatic and grazing controls on ecological properties and processes on the north-eastern Plateau grassland system. Specifically, we are investigating how climate warming, historic and current grazing levels, and the interaction of warming and grazing affect various aspects of rangeland quality. In this paper, we present 1999 growing-season data on habitat, grazing history and treatment effects on vegetative aboveground biomass.
To address our research questions, in 1997 we established experimental plots at the Haibei Alpine Meadow Research Station in Qinghai Province, China (located at 37°29'N, 101°12'E, with an average altitude of 3200 metres above sea level (m.a.s.l.), on the north-eastern portion of the Tibetan Plateau. Annual precipitation averages 500 mm, over 80% of which falls during the summer months. Mean annual air temperature is 2°C (Zhao and Zhou 1999). We are working in two main habitats that are distinguished by the dominant vegetation and the traditional season-of-use by the pastoralists: 1) a summer-grazed, Potentilla fruticosa-dominated deciduous shrubland; and 2) a winter-grazed grassland. The grassland is comprised of a combination of sedges such as Kobresia humilis, perennial grasses such as Elymus nutans and Stipa purpurea and a suite of forbs, over 80% of which are perennial. Within each vegetation type, we identified sites with low and high grazing intensity histories for a total of four main study areas. Grazing histories of the sites over the past 10–15 years were reconstructed based on interviews with local herders whose families have lived in the region for several generations and with long-term researchers at the station.
Within each study area, we fenced off an area of 30 × 30 m, in which we established a complete factorial experimental design with four-fold replications. The treatments are: 1) warming; 2) defoliation; and 3) combined warming and defoliation (Figure 1). The heating treatment is achieved by setting up conical fibreglass open top chambers (OTCs), which are 1.5 m diameter and 40 cm in height. The fibreglass material has a high solar transmittance (86%) in the visible wavelengths and a low transmittance (<5%) in infrared radiation (Marion 1996). OTCs are the standard warming devices used by the International Tundra Experiment Program (ITEX) and are often used in remote locations to simulate climate warming (Marion et al. 1997; Oechel et al. 1998). Clipping plots to a height of 1–2 cm aboveground simulates grazing, while hand plucking shrub leaves simulate browsing. The grazing treatments at all sites simulate the amount of biomass removed by herbivores in the high grazed sites outside of the enclosed areas.
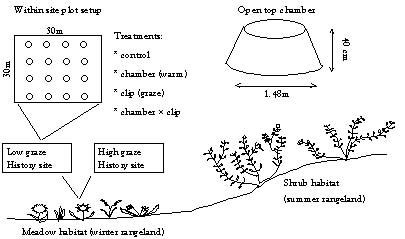
Figure 1. Experimental design.
We report on habitat, grazing history and treatment effects on air temperature and aboveground peak biomass in the plots. In each plot we measure air temperature every hour over the growing season with HoboPro data loggers. Temperature probes are shaded from direct sunlight exposure and are situated within 15 cm of the centre of the plots at 10 cm aboveground. Vegetation sampling occurs in a 75 × 75 cm grid cantered in the plots. Non-destructive aboveground biomass sampling is achieved by measuring areal coverage every three weeks. Regression relationships between areal coverage and harvested aboveground dry biomass (B) off-plot in both habitats have been developed1. Peak biomass occurs when the converted areal coverage values reach a maximum over our sampling period; for the clipped plots, we add the removed biomass to the maximum measured values. Our primary statistical tool is analysis of variance (ANOVA) using SYSTAT version 8.0.
1. Shrub biomass (B) = –2.559 + 1.206 * Areal coverage (AC) (adjusted R2 = 0.941, P<0.0005); Meadow grass B = –4.134 + 1.815 * AC (adjusted R2 = 0.954, P<0.0005); Shrub grass B = 11.672 + 1.177 * AC (adjusted R2 = 0.706, P = 0.006); Forb B = 0.566 + 0.673 * AC (adjusted R2 = 0.829, P<0.0005); Sedge B = 0.269 + 1.248 * AC (adjusted R2 = 0.463, P = 0.12).
In this paper, we refer to the four study sites as follows:
We refer to the grazing history sites as follows:
We refer to the treatments as follows:
Habitat and grazing history control plot site comparisons: The control plots in the meadow sites were 0.80°C warmer (P = 0.13) than the control plots in the shrub sites. The controls in the HGSH site were 0.76°C warmer (P = 0.15) than those in the LGSH site.
Treatment effects: Combining all of the sites (Figure 2), the growing-season-averaged air temperature in the OTC and the OTC × G plots were 1.28°C (P<0.0005) and 1.33°C (P<0.0005) warmer than the controls, respectively. However, there was some variation in how air temperature at the individual sites responded to the treatments (Table 1).
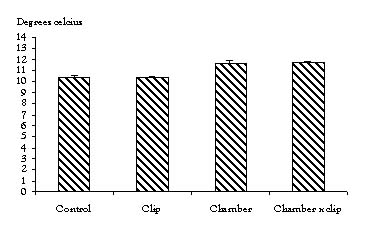
Figure 2. The 1999 growing-season-averaged air temperature with all sites combined.
Table 1. Treatment effects on season-averaged air temperatures in the four study sites.
Study sites |
C |
G |
OTC |
OTC × G |
HGM |
10.16 ± 0.38 |
10.13 ± 0.07 |
12.34 ± 0.10 |
11.43 ± 0.22 |
LGM |
11.08 ± 0.04 |
10.19 ± 0.03 |
11.09 ± 0.09 |
11.95 ± 0.05 |
HGSH |
10.58 ± 0.28 |
10.66 ± 0.09 |
12.55 ± 0.06 |
12.22 ± 0.12 |
LGSH |
9.86 ± 0.15 |
10.56 ± 0.14 |
10.61 ± 0.01 |
11.39 ± 0.16 |
Habitat and grazing history control plot site comparisons: The total peak aboveground biomass values in the control plots of the four sites are illustrated in Figure 3. The control plots in the meadow sites have 94 g/m2 more total biomass (P<0.0005) as compared to the shrub sites. The LGM has 61 g/m2 more total biomass (P = 0.009) as compared to the HGM; conversely, the LGSH has 38 g/m2 less total biomass (P = 0.003) as compared to the HGSH.
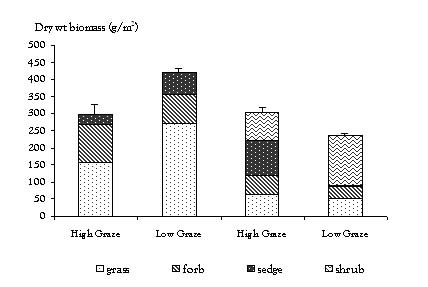
Figure 3. Total and functional group aboveground biomass comparisons among sites
Treatment effects–All sites combined: When all four sites are pooled, the aboveground peak biomass in the chambered plots (OTC and OTC × G treatments combined) was 12% lower (P = 0.07) than in the non-chambered plots (C and G treatments combined) (Figure 4). The biomass in the defoliated plots (G and OTC × G treatments combined) was not significantly different from that in the non-defoliated plots (C and OTC treatments combined). This suggests that when all sites are pooled, the chamber-induced effects of warming are greater than the defoliation effects of grazing with respect to total peak aboveground biomass.
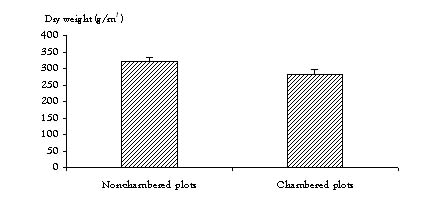
Figure 4. Effects of chamber treatments on total aboveground biomass with all sites combined.
Grazing history comparisons: Different trends emerge when the sites are grouped by grazing history (Figure 5). In both high grazing history sites, there was a significant effect on total biomass of the chamber treatments (OTC and OTC × G plots combined) as compared to the non-chambered treatments (C and G plots combined). The chambered plots in the HGSH had 62 g/m2 less biomass (P = 0.01) and in the HGM had 62 g/m2 less biomass (P = 0.13) as compared to the non-chambered plots. However, in both HG sites, the combined non-defoliated plots (C and OTC treatments) were not significantly different from the defoliated plots (G and OTC × G treatments). In the low grazing history sites, the defoliated treatments had significantly more biomass as compared to the non-defoliated treatments. The defoliated plots in the LGSH had 58 g/m2 more biomass (P = 0.001) and in the LGM had 23 g/m2 more biomass (P = 0.12) as compared to the non-defoliated plots. However, there was no significant effect of the warming treatments on total biomass in the LG history sites. In both grazing history sites, the response of the shrub sites was greater in both magnitude and statistical significance than in the meadow sites.
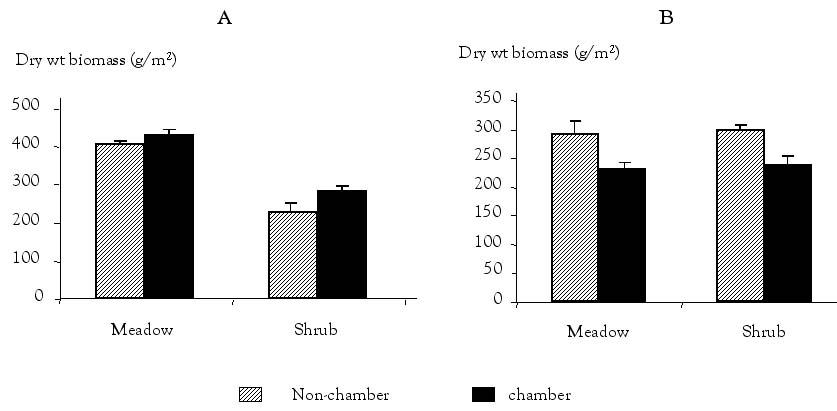
Figure 5. A. Low grazing history sites (LGSH)—effects of defoliation (G and OTC x G plots combined) and non-defoliation (C and OTC plots combined) on total aboveground biomass. B. High grazing history sites (HGSH)—effects of chambers (OTC and OTC x G plots combined) and non-chambers (C and G plots combined) on total aboveground biomass.
Habitat and grazing history control plot site comparisons: Figure 3 illustrates the amount and relative proportions of peak forb, grass, sedge and shrub biomass in the control plots of all four sites. The meadow sites have 162 g/m2 (P<0.0005) more grass biomass and 51 g/m2 (P<0.0005) more forb biomass than the shrub sites. The combined LG sites have 46.37 g/m2 (P = 0.005) more grass biomass, 24.78 g/m2 (P<0.0005) more forb biomass and 31.19 g/m2 (P = 0.001) less sedge biomass than the HG sites.
Forbs: One consistent result across all sites was that forb biomass increased with the defoliation treatments, not considering if they were warmed or not. When all sites are pooled, the combined defoliated plots had 29% (P = 0.01) more forb biomass than the non-defoliated plots (Figure 6).
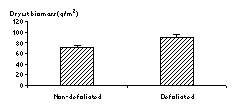
Figure 6. Effects of defoliation on forb biomass with all sites combined.
Grasses: When there was a response of grass biomass to the treatments, the defoliation treatments stimulated grass biomass while the chamber treatments depressed grass biomass. In both shrub sites, the grass biomass was significantly lower in the OTC plots as compared to their respective controls (30%, P = 0.11 decline in the HGSH; 73%, P = 0.001 decline in the LGSH). In the HGSH site, the G treatment resulted in 66% more grass biomass than the C treatment (P = 0.016). The OTC × G treatment resulted in 62% (P<0.0005) more grass biomass than the controls and 131% (P<0.0005) more grass biomass than the OTC treatment. In both LG sites (meadow and shrub sites), the G plots had more grass biomass than the OTC × G plots (18%, P = 0.05 more in the LGM; 70%, P = 0.004 more in the LGSH).
Sedges: In the HGSH and the LGM sites (the two sites with the greatest amount of sedge biomass), the chamber treatments each resulted in significantly less sedge biomass as compared to the controls (HGSH 70%, P<0.0005 decline; LGM 38%, P = 0.05 decline).
Shrubs: The relatively consistent patterns noted above—negative biomass responses to the chambers and positive biomass responses to the defoliation treatments—do not hold with respect to shrub biomass. In the HGSH site, the G plot had 53%, P = 0.01 less shrub biomass than the C plots and 60%, P = 0.001 less biomass than the OTC plots. Moreover, the OTC × G plots in the LGSH had 53%, P = 0.002 more shrub biomass than the controls and 51%, P = 0.003 more biomass than the OTC plots. The OTC×G plots in the HGSH sites had 25%, P = 0.05 less shrub biomass than the controls and 36%, P = 0.018 less shrub biomass than the OTC plots.
Habitat and grazing history control plot site comparisons: At 10 cm aboveground, the valley bottom meadow sites are marginally warmer than the shrub sites, which are approximately 40 m higher in elevation than the meadows and situated on a NW facing slope with moister soils. The HGSH sites were marginally warmer than the LGSH sites. This could be due to the higher cover of shrubs at the LGSH which tend to keep the temperatures under its canopy cooler than those not under the shrub canopy.
Grazing history: The air temperature in the HG sites responded more strongly to the OTC treatments as compared to the LG sites. The HGM has both less total peak biomass and drier soils as compared to the LGM. Moreover, the higher cover and amount of shrub biomass in the LGSH may partially mitigate the amount of warming experienced 10 cm aboveground and maintain moister soils as compared to the HGSH site.
Within sites: Within both LG sites, the OTC × G treatments tend to result in slightly higher temperatures than the OTC treatments. At any given time, the amount of standing biomass (maximum converted area coverage values without the addition of the clipped biomass) was lower in the OTC × G plots as compared to the OTC plots. Harte et al. (1995) found that both vegetative biomass and soil moisture status affect the magnitude of warming expressed at different sites in response to warming manipulations.
The above results suggest that current and historic grazing intensity may affect the expression of warming experienced at the sites. For a given downward flux of infrared radiation, the amount of warming may be greater at sites with a higher historic and current amount of grazing. Since our results indicate that warming tends to further reduce the biomass in these HG sites, this could constitute a positive feedback to warming.
All sites: When all sites are grouped together, the combined chamber treatments resulted in less biomass as compared to the non-chambered treatments. This could be due to decreased growth in response to warming and/or reduced soil moisture availability. Alternatively, the warming treatments may have an indirect effect on biomass. In June 1999, the amount of litter accumulated in the chambered plots was significantly greater (65% more; P<0.0005) than that in the non-chambered plots. This increased litter biomass could hinder plant establishment and growth.
Habitat comparisons: In terms of total biomass response to the treatments, the shrub sites were more responsive to the chamber and defoliation treatments as compared to the meadow sites. This suggests that the total aboveground biomass of the meadow sites might be more resilient to warming and grazing as compared to the shrub sites. The meadow sites may be less responsive to the defoliation treatments as the timing of our clipping treatments simulates the actual timing of grazing in the winter-grazed meadows; we clip the biomass prior to the onset of the growing season. However, this is a relevant finding in light of the different land tenure and season of use patterns between the meadow and shrub sites. Over the past 10–20 years, the winter-used meadow pastures have been allocated to individual households; fencing of these lands serves to demarcate property boundaries and to exclude other livestock from grazing on these individually held parcels. These changes can result in increased grazing pressures on the summer grazing lands (Williams 1996). This increased grazing pressure would occur in the shrub habitat—the vegetative community that our results suggest may be more vulnerable to the effects of warming and grazing.
Grazing history comparisons: In both low grazing history sites (meadow and shrub), the defoliation treatments result in more total aboveground peak biomass than the non-defoliated treatments. This could be evidence of biomass overcompensation, at least in the short-term. Dyer et al. (1993); Turner et al. (1993) reported that grazing increases, decreases or has no net effect on aboveground productivity in other systems. This is a highly controversial area of ecology and rangeland science (Maschinski and Whitham 1989; Painter and Belsky 1993; McNaughton et al. 1997). The outcome depends on a suite of factors, such as climatic conditions, evolutionary history of grazing, and the timing, intensity and frequency of grazing (Milchunas and Lauenroth 1993; Turner et al. 1993). Our short-term (< 5 years) results suggest that when grazing is removed, as it is in our control plots, litter tends to build up. The presence of a substantial litter cover in the LG control plots may cause shading, which, in turn, reduces plant performance. Thus, defoliation of the vegetation, including removal of litter (yak and sheep consume senesced vegetation during the winter months), enhances peak total biomass relative to non-defoliated plots. Furthermore, the LG sites may have more reserves stored belowground. Thus, defoliated plants can reallocate belowground reserves to increased aboveground growth. The litter present in June 1999 in the control plots of the HG sites (32 g/m2) is substantially lower than that in the LG sites (167 g/m2). This smaller amount of litter may not hinder plant establishment and growth; moreover, plants with a higher grazing intensity history may not have the belowground reserves to overcompensate in response to defoliation.
Thus, our results suggest that, at least in the short-term, some amount of grazing may enhance aboveground biomass on the north-eastern Tibetan rangelands. Eliminating grazing from some sites may result in litter accumulation and reduced biomass production. However, with long-term heavy grazing, the rangelands may lose their ability to overcompensate for grazing. Our results suggest that, in the short-term, the sites with high grazing intensity histories can at least equally compensate for the total biomass removed by grazing.
In the HG sites, the combined chamber treatments elicited a stronger total biomass response than the combined defoliation treatments. In the LG sites, the defoliation treatments elicited a stronger total biomass response than the combined chamber treatments. This finding reveals that the effects of climate warming and grazing depend not only on habitat, but also on the land use history of the site. Sites that have higher grazing histories may be more susceptible to the effects of warming as compared to lower grazing history sites. In our study sites, over the short-term, the warming treatments reduced total biomass as compared to the non-warmed plots. Moreover, sites with a low grazing history may be more responsive to shifts in grazing practices as compared to sites with a high grazing history. In our study sites, over the short-term, the defoliation treatments increased total biomass as compared to the non-defoliated plots.
The observed resilience, with respect to total aboveground biomass, of the meadow habitat to our experimental manipulations may in part be explained by compensatory biomass responses of the different vegetative functional groups in the meadow. For example, in the OTC × G plots of the LGM site, forbs increased while sedges decreased by a similar amount. Several studies that have examined climate warming effects on vegetative biomass have observed this compensatory growth among different functional groups that result in no net change at the total biomass level (Chapin et al. 1995; Harte and Shaw 1995).
A consistent response across all sites was that forb biomass increased with the defoliation treatments. This finding is relevant to the issue of rangeland sustainability on the Tibetan grasslands. Some researchers are concerned that increasing forb biomass may enhance small mammal activity, which can contribute to rangeland degradation (Yang 1992; Ma et al. 2000; Smith and Foggin 2000). Moreover, some forbs are important medicinal plants and contribute to the pastoralist livelihood.
In the LGSH sites, the defoliation treatments increased total biomass relative to the non-defoliated plots. For example, in the LGSH site, the G plot and the OTC × G resulted in significantly more biomass as compared to the controls. This could be evidence of biomass overcompensation, especially under the conditions of warming. However, in relating these biomass changes to rangeland quality changes, we must consider not only the direction and amount of biomass change, but also the composition of that biomass. Most of the increase in total biomass in the LGSH was due to enhanced shrub biomass. In fact, grass biomass decreased in both the OTC and the OTC × G plots. Our analysis reveals a negative correlation between shrub and grass biomass (Pearson correlation coefficient = –0.78). Sheep browse the deciduous shrubs in the region, but the yak and horses do not consume the shrubs as forage. Therefore, an increase in shrubs might be favourable for sheep, but would not benefit yak or horses, especially if it entails a concurrent decrease in grass biomass. Thus, the impact of this compositional change in vegetative biomass depends on the pastoralists' herd composition, which is quite variable from region to region on the north-eastern Plateau (Miller 1999a). Moreover, since the shrubs commence growth and flower earlier in the season than most of the grasses, this shift in compositional biomass may be favourable in providing more abundant forage earlier in the season. However, it might also encourage the pastoralists to move onto the summer-grazed shrublands earlier in the season, with further implications for the shrub habitat. Finally, in the HGSH site, total biomass decreased in response to the OTC treatment due to the significant decreases in graminoid species, clearly a negative effect for the grazing herbivores.
Aboveground biomass is important as the primary source of forage for both domestic and wild animals, as medicinal goods, fencing and other amenities. Moreover, aboveground biomass has important indirect effects on rangeland quality. The vegetation that is not removed by the secondary consumers in the system decomposes and contributes to the soil organic matter pool. The organic matter status of the soil affects both the fertility and water-holding capacity of the soil (Tisdale and Oades 1982). Our future papers will report on the soil organic carbon and nutrient availability status of our study sites and how they respond to our experimental manipulations.
This initial analysis of the effects of climate warming and grazing on vegetative aboveground biomass suggests that the winter-grazed meadow sites may be more resilient to these factors as compared to the summer-grazed shrub sites. This resilience may be partially attributed to compensatory biomass growth of different functional groups that stabilises the total biomass responses to our experimental treatments. Moreover, the meadow resilience may also be partially attributed to the timing of defoliation events in the winter-used meadows. Aggregating the data from all sites, the warming treatments resulted in decreased total aboveground biomass. However, the responses to warming and grazing were strongly mediated by habitat type and grazing history. The sites with a high grazing intensity history may be more responsive to warming, whereas sites with a low grazing intensity history may be more responsive to grazing. Our LG history sites provided evidence of biomass overcompensation in response to defoliation, at least in the short-term. However, it is important to understand the compositional changes in biomass and how they will affect rangeland quality.
This paper represents an initial analysis of data from a more comprehensive study. For example, in this paper, we report on the air temperature effects of our treatments—but we are also monitoring soil moisture and soil temperature in all of our plots. Thus, future analyses will help us to discern whether the results we have reported above are due to the direct air warming effect of the treatments or to other indirect effects of warming—such as alterations in soil moisture availability. In addition, we have measured other outcome variables over multiple growing seasons that will provide further insight into our research questions. Moreover, we have only reported treatment effects on growing-season-averaged air temperatures for 1999. Growing season temperature averages may mask important treatment temperature differences over critical growth periods in the life cycle of the vegetation and of the grazing animals. For example, in May 2000 in the LGM site, the air temperature for the OTC plots were 1.79°C warmer than the controls (P<0.005). However, our season-averaged 1999 air temperature data for the LGM reported above do not show any significant changes in air temperature due to the OTC treatment in that site. Finally, we are currently sampling over a larger area both to examine how representative our study sites are of other sites in the region and to relate to our results to a broader spacio-temporal scale.
Despite the limitations and the ongoing nature of our research, we hope that the preliminary findings reported in this paper will enhance our understanding of ecosystem dynamics in the region and contribute to the debate concerning the health and functioning of the Tibetan rangelands. While many people think of the region as remote and set off in traditional ways, the Tibetan Plateau region is facing contemporary challenges of the modern world—from development pressures to global climate change. It is imperative to work both within and beyond the borders of the Tibetan Plateau to address the driving forces behind these perturbations. However, it is also important to recognise these challenges and to obtain a basic and comprehensive understanding of the ecological dynamics in the region to help maintain the rangelands and pastoral lifestyles in the face of these prevailing changes.
We thank Liu Jianzhen, Tsering Thondup, Jia Hu, Sarah McCarthy, Sue Kloss, Suzanne Lippert and Emily Betts for field assistance. The Haibei Research Station provided logistical support. Financial support was provided by the National Science Foundation (USA).
Chapin F.S., Shaver G., Giblin A., Nadelhoffer K. and Laundre J. 1995. Responses of arctic tundra to experimental and observed changes in climate. Ecology 76(3):694–711.
Cincotta R., Zhang Y. and Zhou X. 1992. Transhumant alpine pastoralism in northeastern Qinghai Province: An evaluation of livestock population response during China's agrarian economic reform. Nomadic Peoples 30:3–25.
Dyer M.I., Turner C.L. and Seasteadt T.R. 1993. Herbivory and its consequences. Ecological Applications 3(1):10–16.
French H.M. and Wang B. 1994. Climate controls on high altitude permafrost, Qinghai-Xizang (Tibet) Plateau, China. Permafrost Periglacial Processes 5:87–100.
Harte J. and Shaw R. 1995. Shifting dominance within a montane vegetation community: Results of a climate warming experiment. Science 267:876–880.
Harte J., Torn M.S., Chang F.R., Feifarek B., Kinzig A, Shaw R. and Shen K. 1995. Global warming and soil microclimate: Results from a meadow warming experiment. Ecological Applications 5:132–150.
Limbach W.E., Davis J.B., Bao B.T., Shi D. and Wang C. 2000. The introduction of sustainable development practices of the Qinghai Livestock Development Project. In: Zheng Du (ed), Formation, evolution, environmental changes and sustainable development on the Tibetan Plateau. Academy Press, Beijing, P.R. China. pp. 509–522.
Ma Y., Lang B., Shi D. and Wang Q. 2000. Establishing a pratacultural system— A strategy for rehabilitation of black soil type deteriorated grassland on the Qinghai-Tibet Plateau. In: Zheng Du (ed), Formation, evolution, environmental changes and sustainable development on the Tibetan Plateau. Academy Press, Beijing, P.R. China. pp. 339–344.
Marion G.M. 1996. Temperature enhancement experiments. In: Molau U. and Molgaard P. (eds), ITEX Manual. Danish Polar Centre. Copenhagen, Denmark pp. 17–20.
Marion G.M., Henry G., Freckman D, Johnstone J., Jones G., Jones M.H., Levesque E., Molau U., Molgaard P., Parsons A.N., Svoboda J. and Virginia R.A. 1997. Open-top designs for manipulating field temperatures in high latitude ecosystems. Global Change Biology 3 (suppl. 1):20–32.
Maschinski J. and Whitam T. 1989. The continuum of plant responses to herbivory: The influence of plant association, nutrient availability and timing. American Naturalist 134(1):1–19.
McNaughton S.J., Banyikwa F.F. and McNaughton M.M. 1997. Promotion of the cycling of diet-enhancing nutrients by African grazers. Science 278:1798–1800.
Milchunas D.G. and Lauenroth W.K. 1993. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63:327–366.
Miller D. 1999a. Nomads of the Tibetan Plateau Rangelands in western China—Part two: Pastoral production practices. Rangelands 21(1):16–19.
Miller D. 1999b. Nomads of the Tibetan Plateau Rangelands in western China—Part three: Pastoral development and future challenges. Rangelands 21(2):17–20.
Oechel W., Vourlitis G., Hastings S. and Bryant P. 1998. The effects of water table manipulation and elevated temperature on net CO2 flux of wet sedge tundra ecosystems. Global Change Biology 4:77–90.
Painter E.L. and Belsky A.J. 1993. Application of herbivore optimization theory to rangelands of the western United States. Ecological Applications 3:2–9.
Smith A.T. and Foggin J.M. 2000. The Plateau pika (Ochotona curzoniae): A keystone species for biodiversity on the Tibetan Plateau. In: Zheng Du (ed), Formation, evolution, environmental changes and sustainable development on the Tibetan Plateau. Academy Press, Beijing, P.R. China. pp. 387–397.
Thompson L., Thompson E.M., Davis M., Bolzan J.F., Dai J., Yao T., Gundestrup N., Wu X., Klein L. and Xie Z. 1989. Holocene–Late Pleistocene climate ice core records from Qinghai–Tibetan Plateau. Science 246:474–477.
Tisdale J.M. and Oades J.M. 1982. Organic matter and water stable aggregates in soils. Journal of Soil Science 33:141–163.
Turner C.L., Seastedt T.R. and Dyer M.I. 1993. Maximization of aboveground grassland production: The role of defoliation frequency, intensity and history. Ecological Applications 3(1):175–186.
Williams Dee Mack. 1996. Grassland enclosures: Catalyst of land degradation in Inner Mongolia. Human Organization 55(3):307–313.
Yang Dingguo. 1992. Degradation and protection of grassland on the Qinghai-Tibet Plateau. In: Erosion, debris flow and environment in mountain regions. Proceedings of the Chengdu Symposium. IAHS. 209. pp. 471–476.
Yao T., Shi Y. and Thompson L.G. 1997. High resolution record of paleo-climate since the little ice age from the Tibetan ice cores. Quaternary International 37:19–23.
Zhao Xinquan and Zhou Xingmin. 1999. Ecological basis of alpine meadow ecosystem management in Tibet: Haibei Alpine Meadow Ecosystem Research Station. Ambio 28(8):642–647.
![]()
![]()
![]()